1.ヒトの遺伝情報-染色体(クロモゾーム)
生物は親から子へ膨大な量の遺伝情報を伝えてゆきます。この遺伝情報は細胞の核内 にある染色体(Chromosome) の中にまとめられています。ヒト細胞の核内には23対の染色体があり、これらのうち22対は母親と父親からそれぞれ1つずつ子供に伝えられたものが対になっていて常染色体 と呼ばれます。常染色体は長いものから順に1番から22番まで番号がついています(21だけは22番より短くなっています)。残り1対の染色体は性染色体 と呼ばれ、子供の性別を決定します。性染色体はXとYがあり、女性はXX,男性はXY の組み合わせを持ちます。受精の際に母親からはXと父親からのXが受精に関与すると子供はXX(女性)、母親のXと父親のYが関与するとXY(男性)の子供が生まれます。
クラインフェルター症候群 では2つのXと1つのYをもつ男性が生まれます(XXY)。またターナー症候群 では1つのXだけでYを持たない女性が生まれます(XO)。常染色体異常:ダウン症候群(Down syndrome) :性決定 SRY という遺伝子が関与しています。SRY遺伝子の塩基配列は性別判定の検査にも利用されています。(例外として染色体のクロスオーバーによってSRYがXに移動するとXXの子孫でも男性的な特徴をもち、また何らかの原因でSRYをYから失うと子孫はXYでも女性的な特徴を持ちます。)
X染色体の不活性化(X
inactivation) X
inactivation と呼びます。X染色体が2個以上あっても1つをのぞき、他はすべて不活性化されます(実際にはX inactivationを逃れる遺伝子もあります)。不活性化されたX染色体は顕微鏡でみるとバール体(Barr
body) と呼ばれるかたまりに見えます。ランダム ですが、一度不活性化されるとその後の細胞分裂でもこの不活性化は保たれます。よって体のある部分では父親由来、他の部分では母親由来のXが不活性化された細胞集団が存在するようになります。Xist という遺伝子がかかわっています。この遺伝子の発現によって片方のXが不活性化されます。三毛猫 があげられます。三毛猫は基本的にメスだけ です。これは白斑(劣性)をつくる遺伝子以外にX染色体上に茶色(優性)と黒(劣性)の毛をつくる遺伝子があり、体表のそれぞれの場所でどちらのX染色体が不活性化されるかでその部分が茶色になるか黒になるかが決まるからです。正常のオスはXを1つしか持たないので茶と黒のモザイクにはなりません。ごくまれにオスの三毛猫も生まれますがこれはXXY という染色体パターン(ひとのクラインフェルターにあたる)を持っています。
インプリンティング(Genomic
Imprinting) インプリンティングの例 :(1)IGF-II という遺伝子は父親からのもののみ発現し、母親からのものは発現しません(脳内では両方からのコピーが発現しています) 。プレイダー・ウイリィー症候群(Prader-Willi syndrome)とエンジェルマン症候群(Angelman
syndrome) :この2つの病気は全く異なった症状を示しますが、ともに第15染色体の部分欠損(15q11-13)が関係しています。父親由来の染色体からこの部分が欠損するとプレイダーウイリィー症候群、逆に母親由来の染色体部分が欠損するとエンジェルマン症候群になります。
2.DNA(Deoxyribonucleic Acid)
アデニン(A)、グアニン(G)、シトシン(C)、チミン(T) の4つの塩基(デオキシリボ核酸 )からなります。これらの塩基はその構造からアデニンとグアニンはプリン(Purine) 、シトシンとチミンはピリミジン(Pyrimidine) と呼ばれます。タンパク質をコードする遺伝子はこれらの4つの塩基が正しい順番でならんだ構造をとっています。このDNA鎖はひとでは全体で30億塩基対 ありますが、そのほとんどの配列はすでに決定されています。DNAのメチル化(Methylation):
ほ乳類のシトシン(C) の3%はメチル化(CH3が付く)されています。このメチル化はCpG というCとGが連続した場所に多くみられ,CpGの80%ぼどがメチル化されています。ゲノム上でCpGが高密度に存在する場所をCpGアイランド(Island )といい、遺伝子発現の調節、癌、インプリンティング等に関係していると言われています。一般的にCpGアイランドのシトシンはあまりメチル化されておらず、それ以外の部分のCpGはメチル化されています。おもしろいことに酵母、線虫、ドロソフィラ(ハエ)のゲノムは全くメチル化されていません。通常の塩基配列決定法ではメチル化は検出できないので、メチル化に敏感な制限酵素や化学的な方法で検出する必要があります。1塩基多型性(Single
nucleotide polymorphism:SNP) と呼びます。SNPはひと個人間の薬の効き方の違い等に関係していると言われています。また、乳ガンに関係した遺伝子BRCA1にはこのメチル化部位が変異を起こしている例が報告されています。二本鎖DNA : DNAの鎖は2本がらせん階段のようにペアになった構造をしています。これをダブルへリックス(Double
Helix) といいます。この際、アデニン(A)とチミン(T), シトシン(C)とグアニン(G) はお互いに対になって結合しています。これを塩基の相補性(complimentary) といいます。10個 で1回転します。アデニンとチミンの間は2つ、シトシンとグアニン間には3つの水素結合があり、二本鎖構造を保っています。DNAはマイナスに帯電 しているのはこのリン酸基のためです。
DNA フィンガープリンティング(DNA
Fingerprinting) 反復配列(リピート) 、たとえばVNTR(Variable
Numver of Tandem Repeat :15−70塩基程度の長さ)やSTR(Short Tandem Repeat :2-4塩基)なども個人間でそのリピートの数に大きな変異があります。これらの遺伝情報の違いは最初、DNAを制限酵素で切断するRFLP:Restriction
Fragment Length Polymorphism 法によって検出されていましたが、最近は、PCR(Polymerase Chain Reaction) という方法を使って少量のサンプルから比較的簡単に反復数を検出することができるSTR がよく使われます。これらの変異情報は子孫に遺伝するので、適当な場所や組み合わせを選ぶと個人や親子鑑別を行うことができます。
3.遺伝子の構造と転写
体を構成するタンパク質は遺伝子上にコードされています。ひとの30億塩基対のゲノムDNA上には約数万の遺伝子が存在すると言われています。遺伝子の長さは数百ベースから数百キロベースまでさまざまです。遺伝子の5’側(DNA鎖のリン酸基が付いている方)にはプロモータ と呼ばれる領域があります。RNAポリメラーゼII が結合します。この酵素はDNA鎖からRNA鎖を合成する酵素でいくつかの蛋白の複合体として働きます。RNAポリメラーゼIIはDNAを鋳型にRNAを合成します。遺伝子上にはエクソン とイントロン と呼ばれる領域があり、イントロン部分はGT・・・AG というコンセンサスで囲まれています。このイントロン部分が削られてメッセンジャーRNA(mRNA) になります。このようにDNAからmRNAが作られることを転写(Transcription) といいます。mRNAの3’側(水酸基がある方)には数十から数百ベースのアデニン(A)が付加され、ポリAテイル(PolyA
tail) を構成します。RNA合成にはDNAのチミン(T)の代わりにウラシル(U) が使われます。
4.コドンと翻訳、蛋白合成
翻訳(Translation) といいます。U(T)、C、A、Gの4つのリボ核酸の組み合わせはアミン酸をコードしています。これは3つごとの塩基のセットが1つのアミン酸に対応していますこれをコドン(Codon) といいます。。たとえばATGはメチオニン(M)、GAAはグルタミン酸(E)に対応しています。4種類の塩基を3つ組み合わせると(4種類X4種類X4種類=64種類)のアミノ酸をコードできますが、実際に蛋白を構成するアミノ酸は20種類 しかなくいくつかのアミン酸のコードは重複 しています(コドン対応表参照)。たとえばアルギニン(R)をコードするコドンは6つあります。アミノ酸以外に翻訳をストップさせるコードはUAA、UAG、UGAの3種類 あります。メチオニン(M) から始まります。このペプチド鎖の合成はトランスファーRNA(tRNA) によって行われます。アンチコドン を持っています。このアンチコドンがmRNA上のコドン配列を認識して対応するアミノ酸を順に結合していきます。スタートコドンからはじまった翻訳はストップコドンまで達すると蛋白合成がストップします。これら蛋白合成は細胞質中にあるリボゾーム(Ribosome) で行われます。DNA → RNA → 蛋白 の方向に伝わります(セントラルドグマ) 。
「ビジュアル生理学」のTOPへ
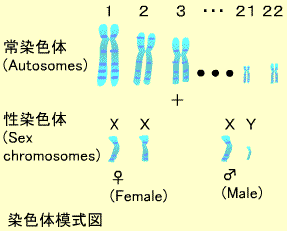
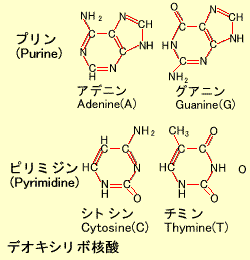 染色体は巨大な糸巻きのようなもので、ヒストンというタンパク質のまわりにDNA鎖が巻き付いています。DNAはアデニン(A)、グアニン(G)、シトシン(C)、チミン(T)の4つの塩基(デオキシリボ核酸)からなります。これらの塩基はその構造からアデニンとグアニンはプリン(Purine)、シトシンとチミンはピリミジン(Pyrimidine)と呼ばれます。タンパク質をコードする遺伝子はこれらの4つの塩基が正しい順番でならんだ構造をとっています。このDNA鎖はひとでは全体で30億塩基対ありますが、そのほとんどの配列はすでに決定されています。
染色体は巨大な糸巻きのようなもので、ヒストンというタンパク質のまわりにDNA鎖が巻き付いています。DNAはアデニン(A)、グアニン(G)、シトシン(C)、チミン(T)の4つの塩基(デオキシリボ核酸)からなります。これらの塩基はその構造からアデニンとグアニンはプリン(Purine)、シトシンとチミンはピリミジン(Pyrimidine)と呼ばれます。タンパク質をコードする遺伝子はこれらの4つの塩基が正しい順番でならんだ構造をとっています。このDNA鎖はひとでは全体で30億塩基対ありますが、そのほとんどの配列はすでに決定されています。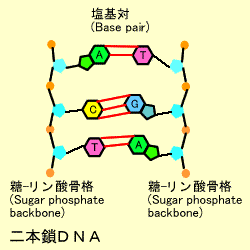
 染色体DNAの特定の部分を比べると個人によって違いのある配列があります。また染色体上に多数存在する反復配列(リピート)、たとえばVNTR(Variable
Numver of Tandem Repeat:15−70塩基程度の長さ)やSTR(Short Tandem Repeat:2-4塩基)なども個人間でそのリピートの数に大きな変異があります。これらの遺伝情報の違いは最初、DNAを制限酵素で切断するRFLP:Restriction
Fragment Length Polymorphism法によって検出されていましたが、最近は、PCR(Polymerase Chain Reaction)という方法を使って少量のサンプルから比較的簡単に反復数を検出することができるSTRがよく使われます。これらの変異情報は子孫に遺伝するので、適当な場所や組み合わせを選ぶと個人や親子鑑別を行うことができます。
染色体DNAの特定の部分を比べると個人によって違いのある配列があります。また染色体上に多数存在する反復配列(リピート)、たとえばVNTR(Variable
Numver of Tandem Repeat:15−70塩基程度の長さ)やSTR(Short Tandem Repeat:2-4塩基)なども個人間でそのリピートの数に大きな変異があります。これらの遺伝情報の違いは最初、DNAを制限酵素で切断するRFLP:Restriction
Fragment Length Polymorphism法によって検出されていましたが、最近は、PCR(Polymerase Chain Reaction)という方法を使って少量のサンプルから比較的簡単に反復数を検出することができるSTRがよく使われます。これらの変異情報は子孫に遺伝するので、適当な場所や組み合わせを選ぶと個人や親子鑑別を行うことができます。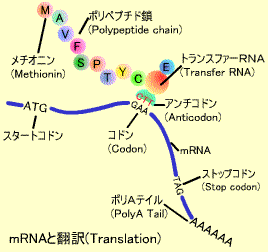 mRNA上のA, U, G, Cの配列はアミノ酸の鎖になり、蛋白として合成されます。これを翻訳(Translation)といいます。U(T)、C、A、Gの4つのリボ核酸の組み合わせはアミン酸をコードしています。これは3つごとの塩基のセットが1つのアミン酸に対応していますこれをコドン(Codon)といいます。。たとえばATGはメチオニン(M)、GAAはグルタミン酸(E)に対応しています。4種類の塩基を3つ組み合わせると(4種類X4種類X4種類=64種類)のアミノ酸をコードできますが、実際に蛋白を構成するアミノ酸は20種類しかなくいくつかのアミン酸のコードは重複しています(コドン対応表参照)。たとえばアルギニン(R)をコードするコドンは6つあります。アミノ酸以外に翻訳をストップさせるコードはUAA、UAG、UGAの3種類あります。
mRNA上のA, U, G, Cの配列はアミノ酸の鎖になり、蛋白として合成されます。これを翻訳(Translation)といいます。U(T)、C、A、Gの4つのリボ核酸の組み合わせはアミン酸をコードしています。これは3つごとの塩基のセットが1つのアミン酸に対応していますこれをコドン(Codon)といいます。。たとえばATGはメチオニン(M)、GAAはグルタミン酸(E)に対応しています。4種類の塩基を3つ組み合わせると(4種類X4種類X4種類=64種類)のアミノ酸をコードできますが、実際に蛋白を構成するアミノ酸は20種類しかなくいくつかのアミン酸のコードは重複しています(コドン対応表参照)。たとえばアルギニン(R)をコードするコドンは6つあります。アミノ酸以外に翻訳をストップさせるコードはUAA、UAG、UGAの3種類あります。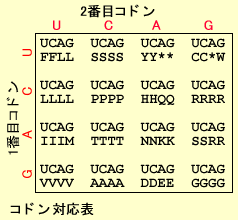 20種類のそれぞれのアミノ酸をもったtRNAはコドンと相補的なアンチコドンを持っています。このアンチコドンがmRNA上のコドン配列を認識して対応するアミノ酸を順に結合していきます。スタートコドンからはじまった翻訳はストップコドンまで達すると蛋白合成がストップします。これら蛋白合成は細胞質中にあるリボゾーム(Ribosome)で行われます。
20種類のそれぞれのアミノ酸をもったtRNAはコドンと相補的なアンチコドンを持っています。このアンチコドンがmRNA上のコドン配列を認識して対応するアミノ酸を順に結合していきます。スタートコドンからはじまった翻訳はストップコドンまで達すると蛋白合成がストップします。これら蛋白合成は細胞質中にあるリボゾーム(Ribosome)で行われます。